| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Природа мембранного потенциала покоя. 12
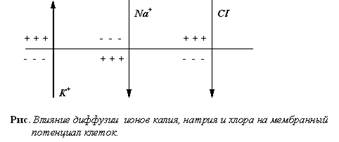
Потенциал покоя – это разность между электрическими потенциалами внутри и вне клетки в состоянии покоя. Величина ПП варьируется в пределах 30-90 мВ. 1. Мембранные потенциалы и их ионная природа Мембранная теория биопотенциалов была выдвинута еще в 1902 году Бернштейном. Но только в 50-х годах эта теория была по-настоящему развита и экспериментально обоснована Ходжкиным, которому принадлежат основные идеи и теории о роли ионных градиентов в возникновении биопотенциалов и о механизме распределения ионов между клеткой и средой.Сущность этой теории заключается в том, что потенциал покоя и потенциал действия являются по своей природе мембранными потенциалами, обусловленными полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и средой, которое поддерживается механизмами активного переноса, локализованными в самой мембране. 1.1 Потенциал покоя, уравнение Нернста Между внутренней и наружной поверхностями клеточной мембраны всегда существует разность электрических потенциалов. Эта разность потенциалов, измеренная в состоянии физиологического покоя клетки, называется потенциалом покоя.Причиной возникновения потенциалов клеток как в покое, так и при возбуждении является неравномерное распределение ионов калия и натрия между содержимым клеток и окружающей средой. Концентрация ионов калия внутри клеток в 20 - 40 раз превышает их содержание в окружающей клетку жидкости. Напротив, концентрация натрия в межклеточной жидкости в 10 - 20 раз выше, чем внутри клеток. Такое неравномерное распределение ионов обусловлено активным переносом ионов - работой натрий-калиевого насоса.Как было установлено, возникновение потенциала покоя обусловлено, в основном, наличием концентрационного градиента ионов калия и неодинаковой проницаемостью клеточных мембран для различных ионов.Согласно теории Ходжкина, Хаксли, Катца, клеточная мембрана в состоянии покоя проницаема, в основном, только для ионов калия. Ионы калия диффундируют по концентрационному градиенту через клеточную мембрану в окружающую жидкость; анионы не могут проникать через мембрану и остаются на ее внутренней стороне. Так как ионы калия имеют положительный заряд, а анионы, остающиеся на внутренней поверхности мембраны, - отрицательный, то внешняя поверхность мембраны при этом заряжается положительно, а внутренняя - отрицательно.
Понятно, что диффузия продолжается только до того момента, пока не установится равновесие между силами, возникающего электрического поля и силами диффузии. Если принять, что потенциал покоя определяется диффузией только ионов калия из цитоплазмы наружу, то его величина E может быть найдена из уравнения Нернста: мембранный потенциал клетка электродиффузия
где [K]i и [K]e - активность ионов калия внутри и снаружи клетки; F - число Фародея;T - абсолютная температура; E - изменение потенциала; R - газовая константа. 7.Условия возникновения ПД. Закон "все или ничего". ПД – это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала покоя вследствие перемещения ионов в клетку и из клетки и способный распространятся без затухания. Выделяют три фазы ПД:
Фаза деполяризации: При действии деполяризующего раздражителя на клетку (например, электрического тока) начальная деполяризация клеточной мембраны происходит без изменения ее проницаемости для ионов. Когда деполяризация достигает примерно 50% порогового потенциала, возрастает проницаемость мембраны клетки для Na+. Условием, обеспечивающим вход Na+ в клетку, является увеличение проницаемости клеточной мембраны, которая определяется состоянием воротного механизма Na–каналов (расположен на внешней и внутренней сторонах клеточной мембраны). Когда деполяризация клетки достигает критической величины – 50 мВ, проницаемость мембраны для Na+ резко возрастает: открывается большое число Na-каналов и Na+ лавиной устремляется в клетку. В результате интенсивного тока Na+ внутрь клетки процесс деполяризации проходит очень быстро. В итоге ПП (потенциал покоя) исчезает, становится равным нулю. Фаза деполяризации на этом заканчивается. 8.Рефрактерность мембраны нейрона: причины возникновения и значение. Потенциал действия (ПД) может распространяться по мембране в обе стороны от места своего первоначального возникновения. Но в физиологических условиях ПД, возникающий, к примеру, в теле нейрона, распространяется на периферию по отросткам только в одном направлении. Обратного распространения ПД не происходит в связи с тем, что в течение некоторого времени тот участок мембраны, в котором развился ПД, становится невозбудимым. Это состояние невозбудимости мембраны называется рефрактерностью. Невозбудимость мембраны обусловлена тем, что после предшествующего возбуждения натриевые каналы на какое-то время становятся инактивированными. Рефрактерностъ — проходящий период состояния мембраны. Причем, если вначале мембрана оказывается нечувствительной к любой силе раздражителя, что обозначается абсолютным рефрактерным периодом, то затем следует относительный рефрактерный период, во время которого надпороговой силы раздражитель уже может вызвать возникновение ПД. Этот период обусловлен постепенным возвратом натриевых каналов к исходному положению, когда закрыты лишь активационные ворота, а инактивационные постепенно открываются.Продолжительность абсолютного рефрактерного периода определяет максимальную частоту генерации потенциалов действия, которая обозначается термином лабильность. Лабильность различных возбудимых структур не одинакова. Она наибольшая в нервных клетках и их отростках, наименьшая в гладкомышечных волокнах. Так, в нейроне абсолютный рефрактерный период около 1 мс, поэтому по нему может проходить до 1000 имп/с. Однако не все нейроны обладают столь высокой лабильностью. Абсолютный рефрактерный период примерно такой же, как и длительность ПД. 9.Активный и пассивный ионный транспорт. Функциональная роль и механизм работы ионных каналов и насосов. ИО́ННЫЕ КАНА́ЛЫ, крупные белковые молекулы и надмолекулярные структуры липопротеидной природы, встроенные в мембраны клетки и ее органоидов (см. Биологические мембраны). Обеспечивают избирательное прохождение ионов через мембрану, в том числе из клетки в наружную среду и обратно. Согласно модели, предложенной в 1972 году Синджером и Николсоном, биологические мембраны имеют жидкостно-мозаичное строение: в жидкой фосфолипидной мембранной пленке «плавают» молекулы белков. В частности, молекулы белков, называемые интегральными, пронизывают мембрану насквозь, выступая одним концом в цитоплазму, а другим — в наружную среду клетки. Внутри такой молекулы белка имеется подобие «дырки», или водной поры, через которую и диффундируют ионы.Трансмембранный транспорт ионов по каналам — основа всех биоэлектрических явлений в организме. Существует большое разнообразие ионных каналов, различающихся по устройству и выполняемым функциям. Численность их может колебаться от нескольких единиц до десятков тысяч на мкм2 мембраны. Простейшими по устройству являются каналы «пассивной утечки» ионов — белковые «дырки» в мембране, пропускающие любые ионы. Они постоянно открыты независимо от действия химических регуляторов или электрического поля. Число таких каналов невелико. Значительно большую и важную группу составляют каналы утечки, избирательно пропускающие один тип ионов (в первую очередь — ионы калия). Избирательный выход калия по концентрационному градиенту из клетки и задержка анионов, не проникающих через мембрану, обусловливает разделение зарядов по разные стороны мембраны и формирование потенциала покоя у любой клетки. Наряду с постоянно открытыми ионными каналами утечки, на мембране любой клетки существуют и другие ионные каналы. Большую часть времени они закрыты и открываются лишь на короткое время и только в ответ на действие особого сигнала: известны потенциалоактивируемые (открывающиеся только в ответ на изменения электрического поля на мембране) и хемоактивируемые (открывающиеся только в ответ на действие определенных химических реагентов) каналы. В отличие от каналов утечки, характерных для любой клетки и участвующих в формировании потенциала покоя клеток, потенциалоактивируемые ионные каналы имеются только у возбудимых клеток — нейронов, мышечных клеток и некоторых других. Именно они принимают непосредственное участие в генерации потенциала действия и возбуждении клетки.Молекулы потенциалоактивируемых каналов имеют сложное устройство; их важнейшие функциональные компоненты — ионселективный фильтр и воротный механизм канала. Селективные фильтры имеют специальные места сужения и особым образом расположенные заряженные химические группировки внутри канала, что позволяет пропускать только один тип иона. Известны потенциалактивируемые каналы, пропускающие только натрий, только калий либо только кальций. Воротный механизм — это группа атомов в молекуле канала, несущая электрический заряд и способная к локальным смещениям (конформациям) внутри молекулы канала в ответ на действие электрического поля. Перемещающиеся группировки, именуемые «воротными системами», в зависимости от своего местоположения либо перекрывают просвет ионной поры, то есть запирают (инактивируют) канал, либо открывают просвет поры. Причем в канале сосуществуют два «воротных» механизма, действующие в противоположном направлении. В покое, когда канал закрыт, одни «ворота» (активационные) закрыты, а инактивационные — открыты. При действии раздражающего электрического стимула (деполяризации) обе воротные заряженные группировки смещаются в канале почти одновременно, но в противоположном направлении. Поскольку инактивационные ворота смещаются более медленно, канал успевает сначала активироваться (открыться), а затем — инактивироваться (закрыться). Присутствие двух типов «ворот» в составе канала обеспечивает возможность саморегуляции работы канала.Хемоактивируемый тип ионных каналов открывается и пропускает ионы только после взаимодействия с соответствующими химическими реагентами — гормонами, медиаторами и т. п. (например, ацетилхолином, адреналином, гистамином). Для этого каналы имеют на своей наружной либо внутренней (цитоплазматической) поверхности специальные активные центры.Еще один тип ионных каналов, обнаруженный в 1980-е годы, — это ионные каналы, чувствительные к растяжению. Они открываются и пропускают ионы в ответ на натяжение мембраны. Встречается у возбудимых и невозбудимых клеток. Отслеживая степень растяжения мышечных органов, такие каналы играют большую роль в поддержании активности клеток сердца, гладкомышечных клеток сосудистой стенки и т. д.Наряду с каналами пассивного транспорта ионов, существует большой класс каналов активного ионного транспорта, называемых ионными насосами. Ионные насосы переносят ионы через мембрану против их концентрационного градиента, используя энергию, выделяемую при гидролизе АТФ. Молекулы ионных насосов — крупные трансмембранные белки (с молекулярной массой порядка 150000), способные связывать и расщеплять АТФ. Например, натрий-калиевый насос осуществляет сопряженный перенос натрия наружу, а калия — внутрь клетки. За 1 секунду этот насос переносит около 200 ионов Na+ из клетки и 130 ионов К+ в клетку. Широко распространен также и Са2+-насос (кальциевая АТФ-аза), откачивающий избыток ионов кальция из клетки. Системы активного ионного транспорта, обеспечивая постоянство концентраций определенных ионов в клетках, играют чрезвычайно важную роль в поддержании уровня потенциала покоя клетки. 11.Типы рецепторов к медиаторам. Понятие об агонистах и антагонистах. Медиаторы (лат. mediator посредник: синоним нейромедиаторы) биологически активные вещества, секретируемые нервными окончаниями и обусловливающие передачу нервных импульсов в синапсах. В качестве М. могут выступать самые различные вещества. Всего насчитывается около 30 видов медиаторов, однако лишь семь из них (ацетилхолин, норадреналин, дофамин, серотонин, гамма-аминомасляную кислоту, глицин и глутаминовую кислоту) принято относить к «классическим» медиаторам. Участие М. в передаче нервного импульса представляется следующим образом. Специализированный для секреции М. участок пресинаптической клетки имеет особую наружную так называемую секреторную мембрану, которая при возбуждении пресинаптической клетки формирует мембранный пузырек, содержащий М. Содержимое пузырька изливается затем в синаптическую щель, диффундирует к постсинаптической мембране, где взаимодействует с ее специфическими рецепторами. При изучении действия М. на рецепторы периферических органов и ц.н.с. выявлены различные типы рецепторов к одному и тому же медиатору (м-, н-холинорецепторы, α-, β-адренорецепторы и др.). Их разделение основано на особенностях биохимических реакций, протекающих в системе медиатор — рецептор. Например, в м-рецепторах реакция носит мускариноподобный характер (они не чувствительны к яду кураре), в н-рецепторах — никотиноподобный (чувствительны к яду кураре). Взаимодействие медиаторов с α-рецепторами вызывает эффект возбуждения (сужение сосудов, сокращение матки и т.д.): с β-рецепторами — тормозные эффекты (расширение сосудов, расслабление бронхов). Вместе с тем α- и β-рецепторы, расположенные в различных органах, могут по-разному реагировать на медиаторы. В зависимости от характера взаимодействия α- и β-рецепторов с различными М. эти рецепторы соответственно разделяют на α1-, α2-, β1- и β2-адренорецепторы. 12 |


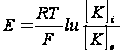 ,
,