| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
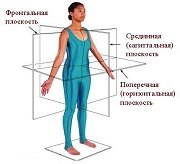
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Биология / Физиология растений / Фотосинтез.
Страница 7 П = Q -R - Тп + Rп где П - поглощенная посевом радиация, Q - суммарная радиация, падающая на посев, R - радиация, отраженная от посева и вышедшая за пределы его верхней границы, Тп - радиация, проникшая к почве, Rп - радиация, отраженная от почвы под растительностью. Коэффициент поглощения энергии ФАР (Qп) посевом определяют делением обеих частей формулы на Q: Qп = П/ Q = 1 - R/Q - Тп/Q +Rп/ Q,где R/ Q -альбедо посева, показывающее, какая доля падающей радиации отражается посевом, Тп/ Q- коэффициент пропускания, показывающий, какая доля падающей радиации (Q) достигает почвы под растительностью, Rп/ Q -альбедо почвы под растительностью. Эффективность фотосинтеза можно характеризовать коэффициентом полезного действия, который определяют по формуле: Е% (КПД) = В.100/А, где А - количество энергии , поступившей за период вегетации на 1 га посева, или энергии, которая была поглощена посевом, в кДж, В - количество энергии, накопившейся в органической массе урожая (биологического или хозяйственного), в кДж. Доказано, что для образования продуктов фотосинтеза при всех благоприятных условиях (орошение, высокая концентрация СО2) солнечная энергия используется только на 2%. В среднем КПД фотосинтеза сельскохозяйственных растений в реальных условиях составляет около 0,5-1% (то есть около 16 кДж/м2 в час ), теоретически же возможно повышение уровня этого показателя до 4-6%. Одной из самых насущных задач, стоящих перед практическим сельскохозяйственным производством, и является повышение эффективности фотосинтеза. Условия, влияющие на интенсивность и продуктивность фотосинтеза. На основные показатели фотосинтеза влияют как внутренние факторы, так и внешние абиотические факторы. К внутренним факторам - то есть к эндогенным механизмам регуляции фотосинтеза у растений - относятся: проводимость листа, фотохимическое лимитирование фотосинтеза, биохимическое лимитирование фотосинтеза, гормональное воздействие, донорно-акцепторные отношения, накопление углеводов, возраст листа или растения. Проводимость листа бывает устьичная и остаточная. Она измеряется в сантиметрах в секунду и является величиной, обратной сопротивлению листа, колеблется от 1 см/с при открытых устьицах до 0,02 см/с при закрытых устьицах, показывает скорость прохождения углекислого газа по тканям листа. Фотохимическое лимитирование фотосинтеза происходит при недостатке поступления энергии из световой фазы фотосинтеза, то есть при недостатке освещенности. Биохимическое лимитирование фотосинтеза определяется недостаточным количеством необходимых для фотосинтеза ферментов, в частности рибулезодифосфаткарбоксилазы/оксигеназы, или недостатком собственно субстрата - рибулезодифосфата. Гормональное воздействие проявляется в действии ингибиторов роста, например, АБК, повышение концентрации которой приводит к закрытию устьиц и к снижению интенсивности фотосинтеза. Донорно-акцепторные отношения проявляются в том, что если у растения уменьшается число акцепторов продуктов фотосинтеза (число клубней, плодов), то интенсивность фотосинтеза снижается , если же уменьшается число доноров продуктов фотосинтеза (листьев) (например в результате повреждения вредителями, искусственная частичная дефолиация), то интенсивность фотосинтеза у оставшихся листьев увеличивается. Накопление углеводов (крахмала) может вызвать снижение интенсивности фотосинтеза, хотя вопрос остается до конца не изученным. Возраст листа (растения) определяет повышение интенсивности фотосинтеза у завершившего рост листа и постепенное снижение интенсивности фотосинтеза у стареющего листа за счет деградации хлоропластов. На показатели фотосинтеза значительно влияют такие абиотические факторы, как: освещенность, температура, водный режим, минеральное питание, Троение хлоропласта - двойная мембрана, отделяющая хлоропласт от цитоплазмы, фотосинтетические мембраны - тилакоиды стромы и тилакоиды гран, наличие участков ДНК, способность к цитоплазматическому наследованию. Внутренние части полости тилакоидов гран и межгранальные тилакоиды - это единая замкнутая фотосинтетическая внутримембранная полость, объединенная в единую фотоэнергетическую систему хлоропласта. Грана хлоропласта состоит из 10-30 тилакоидов, а всего в хлоропласте 100-150 гран, таким образом поверхность фотосинтетических мембран тилакоидов в 10 раз превышает поверхность самого хлоропласта. Особая роль отводится концевым тилакоидам граны, которые, будучи селективным фильтром, предохраняют грану от излишнего облучения или подают сигнал на изменение ориентации оси граны. При оптимальных условиях освещения оси гран обычно направлены радиально к более выпуклой стороне хлоропласта. Функция хлоропласта - осуществление процесса световой фазы фотосинтеза и накопление энергии в виде макроэргических молекул (АТФ и НАДФ восстановленного). Свойства хлоропластов - способность к перемещению внутри клетки под воздействием условий освещенности и концентрации углекислого газа. Передвижение хлоропластов по клетке называется фототаксисом или хемотаксисом хлоропластов в зависимости от причины , вызывающей это передвижение. При умеренном освещении хлоропласты выстраиваются таким образом, чтобы на них попадало максимальное количество света, а при избыточном освещении выстраиваются вдоль падающих солнечных лучей. Такое расположение хлоропластов называется парастрофией . Ночью хлоропласты выстраиваются в положении апострофии . Хромопластыпридают желтую, оранжевую, красную окраску лепесткам, плодам, листьям, так как содержат большое количество специфических каротиноидов, обладающих тем или иным оттенком окраски. Хромопласты функционально дополняют деятельность хлоропластов, кроме того выполняют функцию привлечения насекомых-опылителей, животных-распространителей семян. В состав фотосинтетических мембран (тилакоидов) входят специфические фотосинтетические пигменты - хлорофиллы и каротиноиды - погруженные в эти мембраны. Хлорофиллыделятся на четыре разновидности: а, b, c, d. Это органические соединения, содержащие 4 пиррольных кольца, связанных атомами магния и имеющими зеленую окраску. Отличаются между собой хлорофиллы по молекулярной массе: а - имеет молекулярную массу 893 и включает фитоловый и метиловый остаток, b - имеет молекулярную массу 907 и включает фитоловый и метиловый остаток, с - включает только метиловый остаток, d - имеет молекулярную массу 891 и близок к протохлорофиллу. У высших растений встречаются в основном хлорофиллы а и b, а у водорослей - а и с или а и d. Хлорофилл впервые был выделен в 1818 году, к 1940 году была расшифрована его структура, а в 1960 году осуществили синтез хлорофилла. Хлорофиллы - это сложные эфиры дикарбоновой хлорофиллиновой кислоты с двумя спиртами (фитолом и метанолом). В карбоксильных группах хлорофиллиновой кислоты водород замещен остатками метилового и фитолового спиртов. Наличие в порфириновом ядре хлорофилла коньюгированной по кругу системы десяти двойных связей и магния обуславливает характерный для хлорофилла зеленый цвет. Хлорофиллу а присущ темно-зеленый цвет, а хлорофиллу b - светло-зеленый цвет. Остаток фитола придает хлорофиллу липоидные свойства, то есть он может растворяться в жировых растворителях. Хлорофиллам свойственна флуоресценция - т.е. свойство под влиянием падающего света, в свою очередь, излучать свет, при этом длина волны излучаемого света обычно больше длины волны возбуждающего света. В проходящих лучах цвет хлорофилла - изумрудно-зеленый, а в лучах отраженного света хлорофилл приобретает красный цвет, то есть длина волны, отражаемой хлорофиллом, больше, чем длина волны света, возбуждающего излучение хлорофилла. Хлорофиллы различаются по спектрам поглощения, при этом у хлорофилла bпо сравнению с хлорофиллом а полоса поглощения в красной области спектра несколько смещена в сторону коротковолновых лучей, а в сине-фиолетовой области максимум поглощения смещен в сторону длинноволновых (красных) лучей. В хлоропластах листьев хлорофиллов в три раза больше, чем каротиноидов, а в плодах, лепестках, зернах, корнеплодах - наоборот. Каротиноиды являются непременными спутниками хлорофиллов. Они подразделяются на бескислородные (каротины и ликопины, имеющие оранжевую и красную окраску - общая формула - С40Н56) и окисленные (ксантофиллы - общая формула - С40Н56О2). |