| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
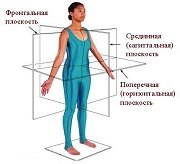
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Костный мозг как периферический лимфоидный орган
В костном мозге происходит продукция протимоцитов и девственных В-лимфоцитов, с одной стороны, и реализация определенных звеньев гуморального и клеточного иммунитета, – с другой. Популяция лимфоидных клеток костного мозга включает незрелые лимфоидные предшественники, также зрелые антитело-продуценты и периферические Т-клетки. Зрелые этапы В-клеточной дифференцировки представлены плазмоцитами (около 1%), которые синтезируют большую часть сывороточных IgG. Для костного мозга характерна длительно продолжающаяся массивная продукция антител на антигены, которые повторно поступают в организм. В популяции лимфоцитов костного мозга присутствуют зрелые Т-клетки: CD3 – 19,8 + /– 4,4%. Отмечается селективный хоуминг зрелых CD8+ лимфоцитов в костный мозг (CD4/CD8 < 1,0). Популяция CD8 клеток костного мозга отличается по иммунофенотипу от CD8 лимфоцитов крови. Практически на всех CD8 лимфоцитах (80-90%) крови и костного мозга присутствуют CD2 и CD3. Фенотип CD4 лимфоцитов крови и костного мозга не различается. Зрелые Т-лимфоциты костного мозга осуществляют контроль над процессами пролиферации и дифференцировки гемопоэтических клеток и клеток костной ткани, как путем контактных взаимодействий, так и опосредованно через продукцию цитокинов. Лимфоидная ткань слизистых (MALT) и кожи Т- и В-клеточные зоны легко идентифицируются в пейеровых бляшках тонкого кишечника, причем Т-клеточная зона расположена непосредственно под эпителием. ВЭВ, через которые лимфоциты с током крови проникают в MALT, расположены в Т-клеточной зоне (так же, как и в лимфатических узлах). На эндотелиальных клетках присутствует молекула адгезии Mad-САМ-1 (апрессин), которая способствует селективному взаимодействию с лимфоцитами пейеровых бляшек, экспрессирующими LPAM-1. Интраэпителиальные лимфоциты экспрессируют специфический антиген HML-1 (CD103), который может быть использован для диагностики лимфом, возникающих из этих клеток. В-клеточные лимфоидные образования кишечника представлены, в основном, разрозненными солитарными фолликулами и пейеровыми бляшками. Эти структуры напоминают фолликулы лимфатических узлов В слизистых и главным образом в коже наиболее часто происходит контакт сенсибилизированных Т-лимфоцитов (хелперные Т-лимфоциты памяти) с чужеродными веществами (презентируются АПК). Одним из проявлений иммунологической реактивности является ГЗТ. ГЗТ может быть вызвана бактериями, вирусами, грибами или изолированными из этих патогенов антигенными препаратами, а также простыми химическими веществами (контактная чувствительность). Эта местная реакция развивается в сенсибилизированном организме и достигает максимума через 24-48 ч после контакта с антигеном. Морфологически представляет собой лимфоцитарно-макрофагальный инфильтрат. В механизме развития реакции главную роль играют Т-хелперные клетки памяти. На ее начальных этапах они распознают антиген в комплексе с собственными молекулами HLA-H на антиген-презентирующих клетках. Активированные Т-хелперные (CD4) клетки продуцируют серию цитокинов и других растворимых факторов (в частности, INFγ и фактор ингибиции миграции), которые способствуют накоплению в ткани макрофагов и их активации, а также стимулируют дифференцировку предшественников Т-цитотоксических клеток в Т-киллеры. Длительное высвобождение цитокинов CD4-лимфоцитами в случаях персистенции антигена ведет к формированию гранулем с накоплением большого количества макрофагов, которые приобретают вид эпителиоидных клеток или сливаются в гигантские и многоядерные клетки. Иммунный ответ Компонентами иммунной системы являются акцессорные клетки: макрофаги и дендритные клетки. Они участвуют в обезвреживании патогенов путем их непосредственного поглощения и уничтожения фагоцитозом, или их превращения в удобоваримую для Т- и В-лимфоцитов форму (этот процесс носит название презентации или представления антигена). Лимфоидные и акцессорные клетки имеют сложную организацию в органах иммунитета. Кооперативные процессы между этими клетками, носящие название адаптивного иммунитета, играют определяющую роль в распознавании чужеродных антигенов и в обеспечении специфичности иммунной реакции в целом. Естественный иммунитет состоит из «неспецифического» внутриклеточного и внеклеточного обезвреживания инфекционных агентов. Внутриклеточное обезвреживание микробов осуществляется профессиональными фагоцитами, к числу которых относятся макрофаги и полиморфноядерные нейтрофилы (микрофаги). Нейтрофилы осуществляют основную защиту против пиогенных бактерий, а макрофаги, наиболее эффективны в отношении бактерий, вирусов и простейших, способных жить в клетках организма-хозяина. В процессе фагоцитоза микроб прикрепляется к поверхности макрофага или нейтрофила, окружается цитоплазматической мембраной и попадает в цитоплазму внутри фагосомы. В течение одной минуты цитоплазматические гранулы нейтрофила или макрофага сливаются с фагосомой и высвобождают свое содержимое (миелопероксидаза, лизоцим, кислые гидролазы) вокруг микроорганизма, разрушая его. Важную роль в обеспечении антимикробного действия фагоцитов играют «кислородный взрыв» (образование супероксидного аниона, активированного кислорода т.п.) и снижение рН в клетке. Убитые микробы перевариваются протеолитическими ферментами, а продукты распада высвобождаются клеткой в окружающую среду. Опсонизация бактерий антителами или СЗb-компонентом комплемента способствует их более эффективному связыванию фагоцитами посредством рецепторов для СЗb и Fc-фрагмента Ig. Внеклеточное разрушение микробов (неспецифическое) осуществляется, главным образом, NK-клетками и эозинофилами. NK-клетки являются большими гранулярными лимфоцитами. Они имеют два типа рецепторов: - лектиноподобные рецепторы, распознающие углеводные структуры и активирующие литический потенциал NK-клетки; - рецепторы, ингибирующие киллинг, блокируют литическое действие NK-клеток в том случае, если оно направлено против клеток, имеющих нормальную структуру молекул HLA-I. Повреждение или делеция молекул HLA-I на клетках-мишенях, распознаваемые рецепторами NК-клеток, способствуют реализации литического действия последних. NK-клетки тесно сближаются с клеткой-мишенью и высвобождают содержимое гранул (гранзимы), что запускает процессы апоптоза в клетках-мишенях. Крупные паразиты, такие, как гельминты, не могут быть фагоцитированы. Они убиваются эозинофилами. В гранулах эозинофилов присутствует главный основной белок, эозинофильный катионный белок, пероксидаза, а также арилсульфатаза В, фосфолипаза D и гистаминаза. На эозинофилах присутствуют рецепторы для СЗb. Активация эозинофилов ведет к мощному респираторному взрыву с образованием активных метаболитов кислорода. Вдобавок, один из белков гранул эозинофилов может разрушать мембрану клетки-мишени. Большинство гельминтов могут альтернативно активировать комплемент, но резистентны к действию С9. Связывание гельминтов с СЗb ведет к последующему прикреплению к ним эозинофилов посредством СЗb-рецепторов. В ходе этой активации эозинофилы запускают механизмы внеклеточной атаки, которые включают высвобождение главного основного белка и особенно катионного белка, повреждающих мембрану паразита. Одной из важнейших Т-клеточных функций является осуществление цитотоксического действия в отношении аутологичных инфицированных клеток. При вирусных инфекциях это касается, в основном, нецитопатических вирусов. CD8 Т-клетки распознают пептиды, происходящие из внутриклеточно синтезированных антигенов, в комплексе с молекулами HLA-I. Основную роль Т-клетки играют в элиминации персистирующих клеток, высвобождающих вирусы, но не гибнущих из-за отсутствия цитопатогенного действия вируса. Основным механизмом цитотоксического действия Т-клеток являются так называемые перфориновая и гранзимная цитотоксичности (последняя индуцирует апоптоз). Важно подчеркнуть две характерных черты Т-клеточного распознавания чужеродных антигенов. 1) Т-хелперные клетки распознают на молекулах HLA-II антигенные пептиды, происходящие из внеклеточных источников. 2) Молекулы HLA-I, которые презентируют антигены CD8 Т-клеткам и экспрессированы почти на всех ядросодержащих клетках, получают пептиды (чаще – вирусные), синтезированные внутриклеточно. Поэтому цитолитическому действию подвергаются только клетки, в которых персистируют живые вирусы, но не клетки, эндоцитировавшие чужеродные белки. В последнем случае Т-хелперный ответ индуцирует гуморальный ответ на внеклеточные антигены (Тн2) или активирует макрофаги.
|