| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
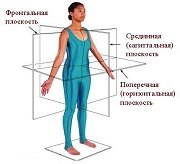
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Полуавтономные органоиды: строение и функции. Теория симбиогенеза. 12
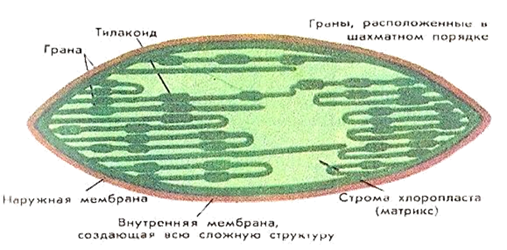
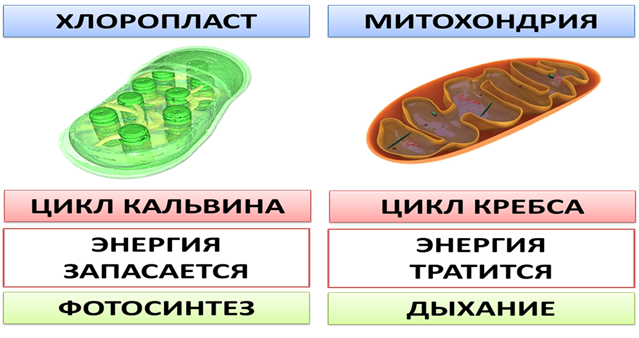
Полуавтономные органоиды- органоиды, содержащие собственные ДНК, все типы РНК, рибосомы, способные синтезировать белки, а также преобразовывать Е в ЕАТФ (синтез АТФ). К таким органоидам относятся двумембранные органоиды: митохондрии и пластиды. Митохондрии и пластиды способны к самовоспроизведению и никогда не образуются из других компонентов клетки. Вне клеток они существовать не могут. Хлоропласты имеют зеленый цвет, обусловленный присутствием основного пигмента — хлорофилла.
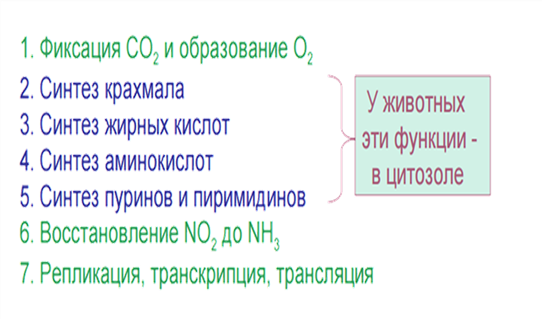
Функции:
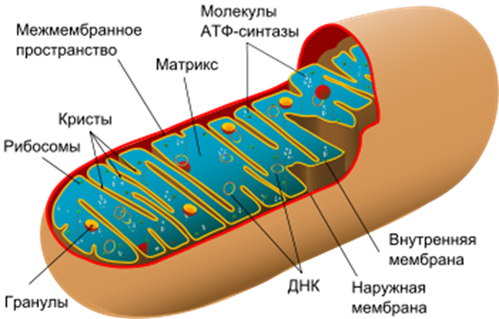
Митохондрия
Возможные пути деления митохондрий: при образовании перегородок или перетяжек Функции:
Теория симбиогенеза согласно этой теории, митохондрии и хлоропласты произошли от симбиотических прокариотических организмов, захваченных протоэукариотом в результате фагоцитоза. Этот протоэукариот, о видимому, представлял собой амебоидный гетеротрофный, анаэробный организм с уже развитыми эукариотическими признаками. Симбиоз большой анаэробной клетки (вероятно, относящейся к архебактериям и сохранившей ферменты гликолитического окисления) с аэробными бактериями оказался взаимовыгодным, причем бактерии со временем утратили самостоятельность и превратились в митохондрии Потеря самостоятельности связана с утратой части генов, которые перешли в хромосомный аппарат клетки-хозяина. Но все же митохондрии сохранили собственный белоксинтезирующий аппарат и способность к размножению. В пользу симбиотического происхождения митохондрий и хлоропластов говорят многие факты. Во-первых, их генетический материал представлен одной кольцевой молекулой ДНК (как и у прокариот), во-вторых, их рибосомы по массе, по строению рРНК и рибосомальных белков близки к таковым у аэробных бактерий и синезеленых. В-третьих, они размножаются как прокариоты и наконец, механизмы белкового синтеза в митохондриях и бактериях чувствительны к одним антибиотикам (стрептомицину), а циклогексимид блокирует синтез белка в цитоплазме. Кроме того, известен один вид амеб, которые не имеют митохондрий и живут в симбиозе с аэробными бактериями, а в клетках некоторых растений обнаружены цианобактерии (синезеленые), сходные по строению с хлоропластами. 15. Ядро: строение и функции. Организация генетического материала эукариотической клетки. Функции ядра 1. Хранение наследственной информации 2. Реализация наследственной информации 3. Воспроизведение молекулы днк 4. Передача информации дочерним клеткам при клеточном делении Геном (совокупность наследственного материала, заключенного в клетке)каждой клетки несет информацию: О первичной структуре всех белков всех клеток всего организма (последовательность аминокислот) О первичной структуре примерно (последовательность нуклеотидов) 60 видов тРНК и различных рРНК (у эукариотов 4 вида) О программе использования данной информации в разных клетках и разных моментах онтогенеза (онтогенез-индивидуальное развитие организма) Состав ядра: Ядерная оболочка Кариоплазма Ядрышко Хроматин или хромосомы (хроматин в неделящемся ядре, хромосомы-в митотическом) Размер: от 1 мкм (простейшие) до 1 мм (в яйцеклетках рыб и земноводных) Оболочка: внутренняя и наружная Наружная соединяется с каналами эпс. На ней располагаются рибосомы. В мембранах ядра имеются поры (3000 - 4000). (проходит обмен веществами между ядром и цитоплазмой) Кариоплазма – желеобразный раствор, заполняет пространство между структурами ядра (хроматином и ядрышками) Ядрышко не окружено мембраной, содержит фибриллярные белковые нити и РНК, исчезают в начале деления клетки и восстанавливаются после его окончания. В ядрышках происходит формирование рибосом, синтез ядерных белков. Образуются на участках перетяжек хромосом. участки хромосом, на которых происходит синтез рибосомных рибонуклеиновых кислот (рРНК Хроматин- деспирализованная форма существования хромосом ГЕТЕРОХРОМАТИН – участки хромосом, находящиеся в конденсированном (упакованном) состоянии в течение всего клеточного цикла. Таким образом, гетерохроматиновые участки в генетическом отношении являются практически неактивными. ЭУХРОМАТИН – основная часть митотических хромосом, в которой локализована большая часть функциональных генов. Эухроматин претерпевает обычный цикл компактизации-декомпактизации во время митоза. Хромосомы- органоиды ядра, являются носителями генов и определяют наследственные свойства клеток и организма.
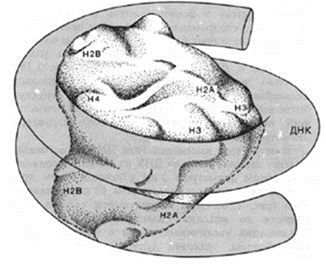
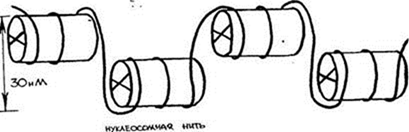
Кариотип- совокупность полного набора хромосом, присущего клеткам данного биологического вида (понятие ввел Левитский в 1924) Кариограмма – систематизированное изображение хромосом, расположенных в ряд по мере убывания их длины. Организация генетического материала эукариотической клетки Длина ДНК диплоидного набора хромосом человека составляет примерно 174 см., средняя длина ДНК одной хромосомы – 5 см. В ядре длина одной хромосомы составляет 0,5 – 1 микрон. Такая упаковка двойной спирали ДНК объясняется ее дальнейшей последовательной компактизацией. 1.Нуклеосомный уровень. Нуклеосома - это ДНК - гистоновый комплекс, который выглядит как частица дисковидной формы диаметром 11 нм. Впервые нуклеосомы были описаны в 1974г. А. Олинс и Д. Олинс. Каждая нуклеосома состоит из белкового кора или октамера и 2 оборотов фрагмента двухцепочечной ДНК
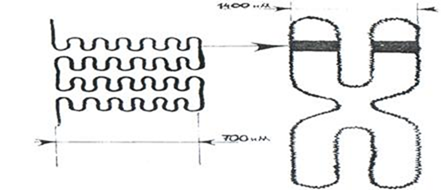
Белковый кор (сердцевина) содержит набор из 4 пар гистоновых белковН2А, Н2В, Н3, Н4. Это самые консервативные белки в любом геноме. Они практически одинаковы у гороха и у человека. Нуклеосомы связываются участками ДНК (линкерная ДНК) свободными от контакта с белковым кором. Укладка линкерного участка ДНК (60-80 п.н.) и соединение нуклеосом друг с другом идут с помощью гистона Н1. Молекула этого белка имеет центральную (глобулярную) часть и вытянутые «плечи». Центральная часть прикрепляется к специфическому участку на поверхности кора, вытянутые «плечи» соединяют соседние нуклеосомы. При этом ДНК наматывается на соседние коры каждый paз в противоположном направлении Выделить нуклеосомы можно непродолжительной обработкой хромосом ферментами дезоксирибонуклеазами. При этом расщепляются участки состыковки нуклеосом. В геноме человека содержатся 1,5 х 107 нуклеосом. Нуклеосомный уровень повышает плотность упаковки ДНК в 7-10 раз. 2. Нуклеомерный уровень. Дальнейшая компактизация ДНК в составе хроматина связана с образованием нуклеосомных комплексов Образуется компактная хроматиновая фибрилла построенная либо по типу соленоида (спиральный тип укладки), либо по нуклеомерному типу (4-12 нуклеосом образуют глобулу).
Нуклеомерная укладка хроматина способствует укорочению нити ДНК примерно в 6 раз, а оба уровня приводят к компактизации ДНК в среднем в 50 раз (42-60). 3. Хромомерный уровень.
Следующий этап компактизации ДНК связан с образованием петлеобразных структур, которые называются хромомерами. При этом возможны два пути упаковки ДНК с помощью негистоновых белков:
Нить нуклеосом разбита на участки по 20 - 80 тысяч пар азотистых оснований (в среднем – 50 тысяч). В местах разбивки находятся молекулы – глобулы - негистоновых хромосомных белков. ДНК - связывающие белки узнают глобулы негистоновых белков и сближают их. Образуется устье петли. Средняя длина петли (300-400 нм) сходна у различных организмов (дрозофила и человек) и включает примерно 50 тысяч оснований. Такую петельную структуру называют интерфазной хромонемой. Хроматин типа «ламповых щеток» - это интерфазный эухроматин. Считают, что петли имеют связи с белками хромосомного каркаса, ядерного матрикса и белками ламины. Укорочение фибриллы на этом уровне происходит в среднем 25 раз, а на всех 3 уровнях в 1000-1500 раз. 4.Хромонемный уровень. При делении клеток идет дальнейшая компактизация хромосом - образование более крупных петель из хромомерной фибриллы. На поверхности упакованные молекулы ДНК несут множество белков, которые образуют подобие чехла. Если удалить этот чехол, то под электронным микроскопом можно отчетливо увидеть, что каждая хроматида построена из хроматиновых петель, отходящих от центральной оси. Диаметр такой упаковки 700 нм
5.Хромосомный уровень. Дальнейшая компактизация хромосом обеспечивается петельной укладкой хромонемной нити, что сокращает их длину примерно в 10 раз.
На этом этапе происходит объединение петель имеющих одинаковую организацию, образуются блоки или минидиски. В образовании одного минидиска участвуют примерно около 20 петель. Таким образом, за счет нескольких уровней компактизации длина ДНК сокращается примерно в 10000 раз. Конденсация хромосом из деконденсированного состояния - это не спирализация, а очень сложный комплекс компактизации, связанный не только с изменением их линейных размеров, но и с регуляцией их работы в процессе жизнедеятельности клетки. Кроме того, компактизация хромосомы - важнейший процесс, связанный с точной передачей наследственной информации очередному поколению. 12 |